高分文献秘籍-细胞毒性T细胞介导的细胞凋亡解决方案
Source: Elabscience®Published: 2025-07-09
树突状细胞(DC细胞)和T细胞相互作用在肿瘤治疗中发挥着关键的作用,有研究表明骨髓来源的DC细胞(BMDCs)诱导CD103+CD8+组织驻留记忆T细胞亚型的产生,可增强生殖道局部肿瘤控制。同时,半乳糖酸阻断进一步提高了BMDCs诱导抗原特异性CD8+ T细胞增殖的能力。全转录组基因表达分析表明,对BMDCs进行仿硅酸处理可诱导参与T细胞活化、细胞粘附和细胞间相互作用的基因差异表达。细胞聚类测定和单细胞嗜性测量表明,糖基化减少的BMDC与CD8+ T细胞形成了更高的嗜性相互作用。DC细胞完成抗原呈递后,在CD8+ T细胞的激活和靶细胞的杀伤过程中,如何发挥协作功能以及调控机能;活化CD8+T细胞的记忆功能分群的变化与其细胞内的乳酸化、棕榈酸化、以及等代谢进程的关系是当前的研究热点之一。这些研究几乎均需要评估CD8+ T细胞的杀伤功能,本期小E将给大家提供DC细胞活化的CD8+ T细胞及评估CD8+ T细胞杀伤功能检测的解决方案。
一、实验原理简介
DC细胞是已知的功能最强抗原呈递细胞(antigen presenting cell,APC),可摄取消化外源抗原并装载在MHC分子中将其呈递给CD4+和CD8+ T细胞以产生免疫反应。共培养DC细胞和CD8+ T细胞是模拟体内CD8+ T细胞激活的最佳方法,因此是CD8+ T细胞相关研究不可替代的模型制备方案。具体而言,表达抗原肽-MHC Ⅰ复合物的DC细胞和CD8+ T细胞共培养时,CD8+ T细胞通过其表面的T细胞受体(TCR)与DC细胞上的抗原肽-MHC Ⅰ复合物结合而被激活。活化的CD8+ T细胞(也称细胞毒性T细胞)可通过溶解介质(如穿孔素和颗粒酶)的释放实现对靶细胞的杀伤力,也可通过T细胞表面Fas配体(FasL/CD95L)与靶细胞Fas受体(CD95)结合,触发Caspase-8激活,直接启动靶细胞的凋亡程序(PMID: 34725504)
二、实验结果展示
1、DC细胞的体外培养和促成熟
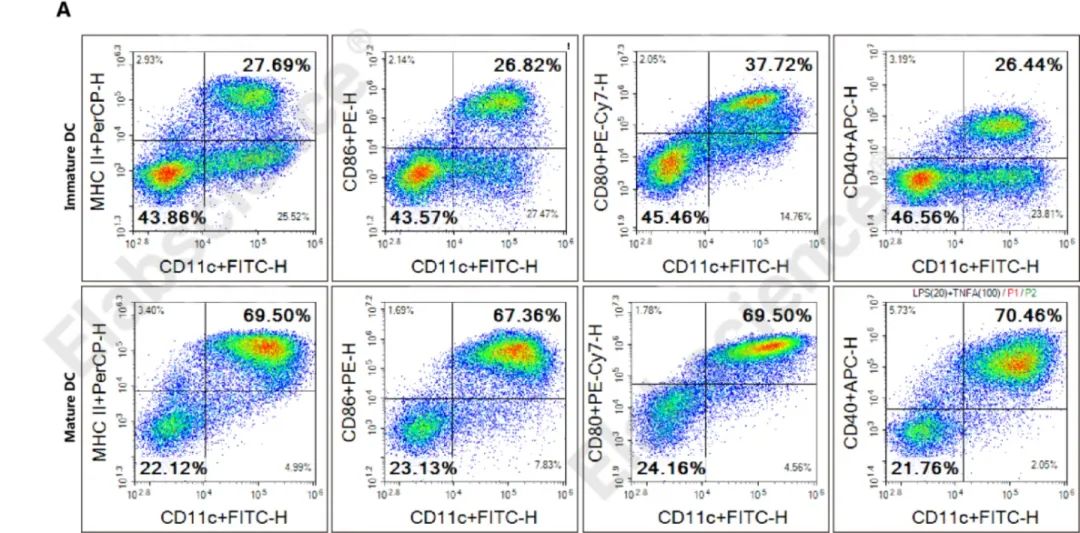
(A)流式细胞术检测体外诱导C57小鼠骨髓细胞为成熟DC细胞的效果,DC细胞经过促成熟和成熟的诱导培养后,MHC II/CD80/CD40/CD86的表达明显升高。
2、CD8+ T细胞的分选和抗原提呈及激活培养
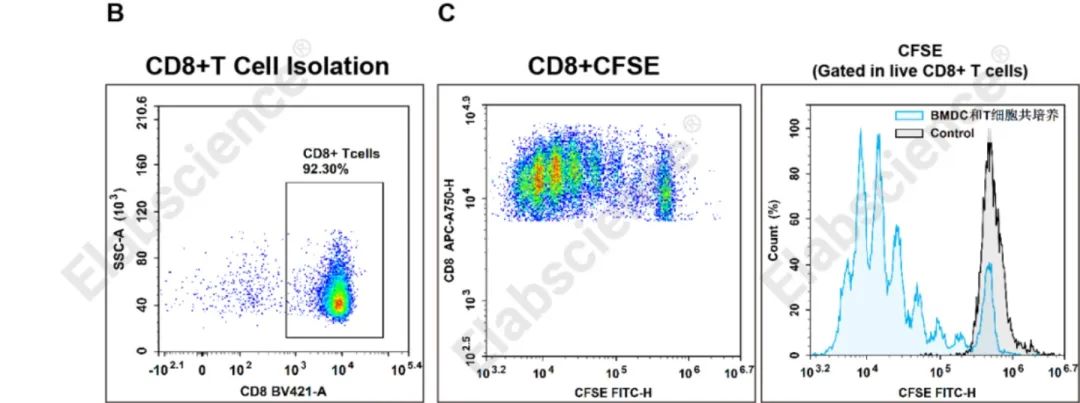
(B)使用小鼠CD8 T细胞阴性分选试剂盒(货号:MIM003N)从C57小鼠脾脏中分选CD8+T细胞,细胞纯度使用流式细胞术进行分析。(C)使用灭活的靶细胞(Raw264.7)抗原装载DC后和分选的CD8+T共培养72 h,检测CD8+T的增殖。
3、CD8+ T细胞杀伤靶细胞的凋亡检测
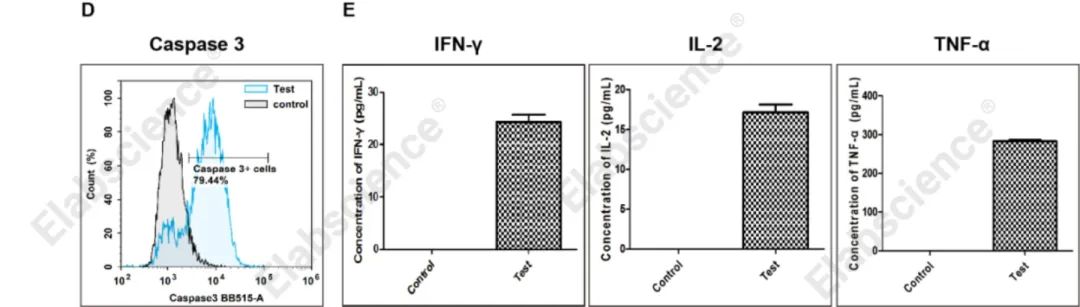
(D)增殖后的T细胞和靶细胞(Raw264.7)按照1:10的比例共培养24 h,流式细胞术检测靶细胞(Raw264.7)中Caspase 3酶活性的变化,和Control组(正常的靶细胞(Raw264.7))相比,共培养组(Test)Raw264.7细胞的Caspase 3酶激活比例增加至79.44%。
(E)ELISA检测细胞培养上清中细胞因子的表达,与Control组(正常的靶细胞(Raw264.7))相比,共培养组(Test)的细胞因子IFN-γ、IL-2、TNF-α表达明显增加。
三、实验方案及操作流程
1、DC细胞的体外培养和促成熟
取小鼠骨髓细胞,使用分化培养基培养6天后,使用成熟培养基培养24 h,获得成熟的DC细胞,详细操作流程请参考”Mouse Bone Marrow-derived Dendritic Cells (BMDC) Induction and Identification Kit”(货号:XJM003)。
80℃水浴加热灭活处理Raw264.7细胞4 h,离心洗涤,使用1640全培养基重悬细胞并计数。(1640全培养基配制方法请参考XJM003说明书)
将灭活处理的Raw264.7细胞和成熟的BMDC按照1:1的比例共培养24 h,收集细胞,100×g低速离心3分钟,去上清,收集细胞沉淀,1640全培养基重悬并计数。
2、CD8+ T细胞的分选和抗原提呈及激活培养
取C57小鼠脾脏细胞,研磨处理,使用EasySort™ Mouse CD8+T Cell Isolation Kit进行细胞分选,检测分选后CD8+T细胞的纯度。
分选后的T细胞加入CFSE染色液标记,37℃染色5 min后终止染色并离心洗涤,使用1640全培养基重悬并计数。
将CFSE标记的T细胞和BMDC细胞按照10:1的比例混合均匀,置入6孔板培养72 h,同时设置单独的CFSE标记的T细胞为Control组(共培养期间每24 h补加二分之一体积的1640全培养基)。
镜下观察,T细胞出现大量的增殖和聚团,收集细胞,150×g离心3 min,使用新鲜的1640全培养基重悬细胞并计数,取少量细胞检测CFSE增殖效果。
3、CD8+ T细胞杀伤靶细胞的凋亡检测
24孔板接种靶细胞(Raw264.7细胞),将T细胞和靶细胞(Raw264.7细胞)按照2:1比例混匀,37℃二氧化碳培养箱中培养24 h后收集细胞沉淀(同时收集细胞上清-20℃冻存备用)并使用PBS洗涤,加入Caspase3流式底物和Elab Fluor® Violet 450 Anti-Mouse CD8a抗体共染,流式上机分析CD8阴性细胞(Raw264.7细胞)群中Caspase 3的酶活情况。(详细的Caspase3检测流程请参考E-CK-A483说明书)
将冻存的细胞上清室温溶解,10000×g离心5 min,收集上清,ELISA检测细胞上清中 IFN-γ、IL-2、 TNF-α的分泌情况。(详细的检测流程请参考ELISA试剂盒E-HSEL-M0007、E-MSEL-M0036、E-HSEL-M0009的说明书流程)。
注:若有更多实验内容方面的细节需要了解,欢迎和我们技术团队沟通,点击联系技术支持!
本实验方案全程使用Elabscience®相关产品,主要产品列于下表:
|
产品名称 |
货号 |
|
小鼠骨髓来源树突状细胞培养和鉴定试剂盒 Mouse Bone Marrow-derived Dendritic Cells (BMDC) Induction and Identification Kit |
|
|
EasySort™-5 磁力架 EasySort™-5 Magnet |
|
|
EasySort™小鼠 CD8+ T 细胞分选试剂盒 EasySort™ Mouse CD8+T Cell Isolation Kit |
|
|
CFSE 细胞增殖示踪试剂盒 CFSE Cell Division Tracker Kit |
|
|
Caspase 3/7 活性检测底物 (流式细胞术用) Caspase 3/7 Activity Detection Substrate for Flow Cytometry |
|
|
FITC 标记抗小鼠 CD3 抗体 [17A2] FITC Anti-Mouse CD3 Antibody[17A2] |
|
|
Elab Fluor®Violet 450 抗小鼠 CD8a 抗体 [53-6.7] Elab Fluor® Violet 450 Anti-Mouse CD8a Antibody [53-6.7] |
|
|
细胞亚铜离子荧光法测试盒 |
|
|
高敏小鼠 IFN-γ (干扰素 γ) ELISA 试剂盒 High Sensitivity Mouse IFN-γ (Interferon Gamma) ELISA Kit |
|
|
微量样本小鼠 IL-2 (白细胞介素 2) ELISA 试剂盒 Mini Sample Mouse IL-2 (Interleukin 2) ELISA Kit |
|
|
高敏小鼠 TNF-α (肿瘤坏死因子 α) ELISA 试剂盒 High Sensitivity Mouse TNF-α (Tumor Necrosis Factor Alpha) ELISA Kit |
四、更多体外模型处理方案参考
细胞凋亡常用的诱导剂以及体外研究的凋亡模型推荐以下:
常用诱导剂和抑制剂
1、常用诱导剂:
星孢菌素(Staurosporine,广谱蛋白激酶抑制剂)
喜树碱(抗肿瘤药物,通过抑制拓扑异构酶活性,干扰 DNA 合成)
常用抑制剂:
Z-VAD-FMK(广谱的 Caspase 抑制剂)
2、细胞凋亡模型
星孢菌素刺激Jurkat /HL-60细胞凋亡模型:5-10 μM星孢菌素诱导3-6 h
喜树碱刺激Jurkat/HL-60/Molt-4细胞凋亡模型:5-10 μM喜树碱诱导3-6 h
顺铂刺激Hela 细胞凋亡模型:3-5 μM顺铂诱导16-24 h
细胞毒性CD8+T细胞杀伤模型:细胞毒性CD8T细胞和目标细胞共培养
更多的铁死亡研究试剂盒、铜死亡研究试剂盒、双硫死亡试剂盒、乳酸化和棕榈酸化研究试剂盒均已上线,若有需要,请随时联系!
参考文献:
[1] Munir S , Hillyer P , Le Nou?N C ,et al.Respiratory Syncytial Virus Interferon Antagonist NS1 Protein Suppresses and Skews the Human T Lymphocyte Response[J].PLoS Pathogens, 2011, 7(4):1-17.DOI:10.1371/journal.ppat.1001336.
[2] Wang C , Barnoud C , Kizil B ,et al.Dendritic Cells Direct Circadian Anti‐Tumor Immune Response[J].FASEB Journal, 2022, 36.DOI:10.1096/fasebj.2022.36.S1.R2866.
[3] Yang B , Kim S , Jung W J ,et al.CTCF controls three-dimensional enhancer network underlying the inflammatory response of bone marrow-derived dendritic cells[J].Nature Communications, 2023, 14(1).DOI:10.1038/s41467-023-36948-5.
[4] Huang Y , Zhou L , Zhang H ,et al.BMDCs induce the generation of the CD103+CD8+ tissue-resident memory T cell subtype, which amplifies local tumor control in the genital tract[J].Cellular Immunology, 2022, 374:104502-.DOI:10.1016/j.cellimm.2022.104502.
[5] Balneger N , Cornelissen L A M , Wassink M ,et al.Sialic acid blockade in dendritic cells enhances CD8+ Tcell responses by facilitating high-avidity interactions[J].Cellular and Molecular Life Sciences, 2022, 79(2):98-.DOI:10.1007/s00018-021-04027-x.
[6] Waldman A D , Fritz J M , Lenardo M J .A guide to cancer immunotherapy: from T cell basic science to clinical practice[J].Nature Reviews Immunology[2025-06-30].DOI:10.1038/s41577-020-0306-5.
[7] Longchao Liu, et al. Rejuvenation of tumour-specific T cells through bispecific antibodies targeting PD-L1 on dendritic cells. Nature Biomedical Engineering. 2021. Doi:10.1038/s41551-021-00800-2.




